Nature | 攀援纤维的 “去抑制” 密码,重新定义小脑学习的核心环路
发表时间:2026-03-30 15:18
今天为大家解读一篇发表于《Nature》的研究论文《Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals》。该研究由哈佛医学院Wade G. Regehr团队完成,完美解决了小脑领域长期存在的经典悖论,把突触解剖、细胞生理、在体环路、功能学习串成完整故事。


研究背景
小脑的运动学习依赖于攀缘纤维(CF) 向浦肯野细胞(PC) 传递的指令信号。CF的输入能诱导PC树突钙信号升高,进而引发长时程抑制(LTD),这是小脑学习的核心机制之一。
然而,一个长期存在的“矛盾”是:CF在兴奋PC的同时,也会兴奋分子层中间神经元(MLI),而MLI中的一类亚型(MLI1)恰恰是抑制PC的,这会抑制树突钙信号,从而阻碍LTD的发生。
那么,CF是如何在不被MLI抑制“干扰”的情况下,有效诱导学习的呢?
研究亮点
1. 结构-功能高度一致:从EM重建到电生理,再到在体记录和钙成像,多尺度验证了CF-MLI2通路的结构基础和功能优势。
2. 解决了长期矛盾:揭示了CF如何通过“去抑制”机制,绕过MLI1的抑制作用,确保LTD所需的钙信号得以增强。
3. 强调同步性的重要性:CF同步激活是打破“抑制-去抑制”平衡、实现有效学习的关键。
4. 模型与实验互补:构建了基于真实解剖数据的计算模型,验证了同步性对环路输出的调控作用。
核心发现
1. MLI存在两种功能相反的亚型
· MLI1:抑制PC,限制钙信号,抑制LTD。
· MLI2:抑制MLI1,从而去抑制(disinhibit)PC,促进钙信号和LTD。
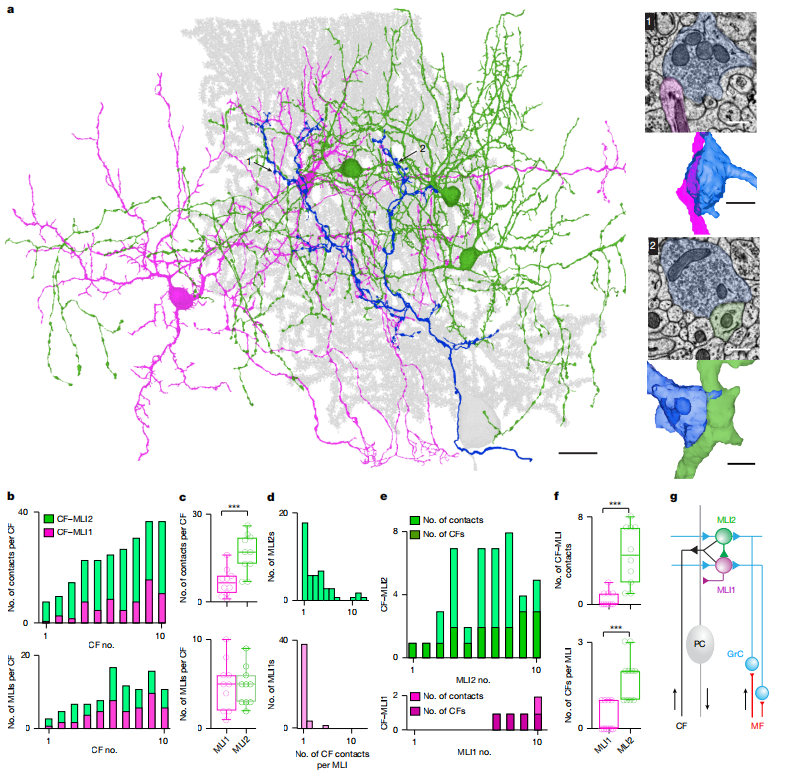
图1
2. CF优先通过“谷氨酸溢出”激活MLI2
通过连续电子显微镜(EM)重建发现:
· CF与MLI2的接触点更多、接触面积更大;
· CF并不形成传统突触结构,而是通过谷氨酸溢出激活MLI2。
·
电生理实验证实:
· CF-MLI2的突触反应更强、更可靠,且包含明显的NMDA成分;
· CF-MLI1的反应则弱得多,且几乎不含NMDA成分。
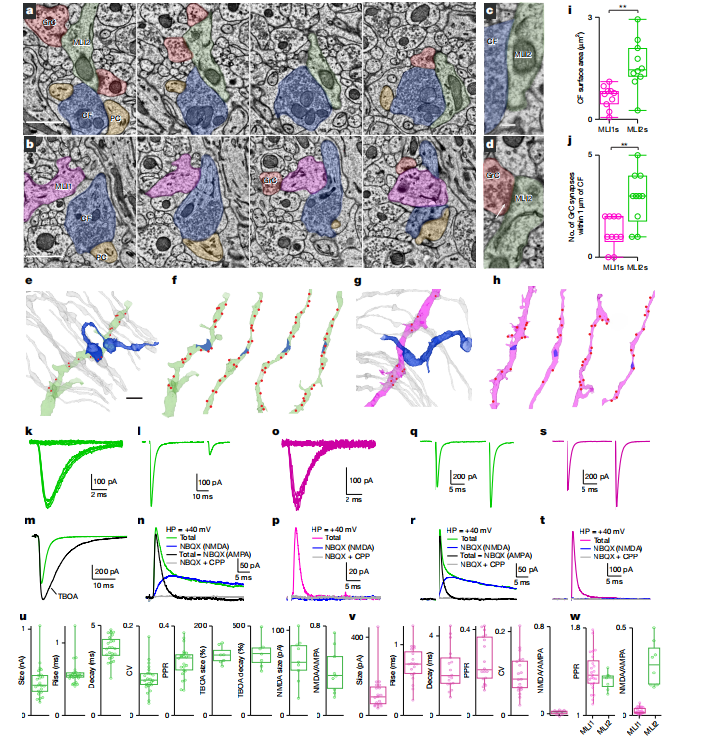
图2
3. 在体记录揭示CF激活的去抑制通路
使用Neuropixels探针在自由运动小鼠中记录:
· CF自发活动优先激活MLI2,抑制MLI1,最终去抑制PC。
·
通过交叉相关分析,明确了信号传递的时间序列:
· CF → MLI2(~0.5 ms)→ MLI1抑制(~2.4 ms)→ PC去抑制(~4.1 ms)
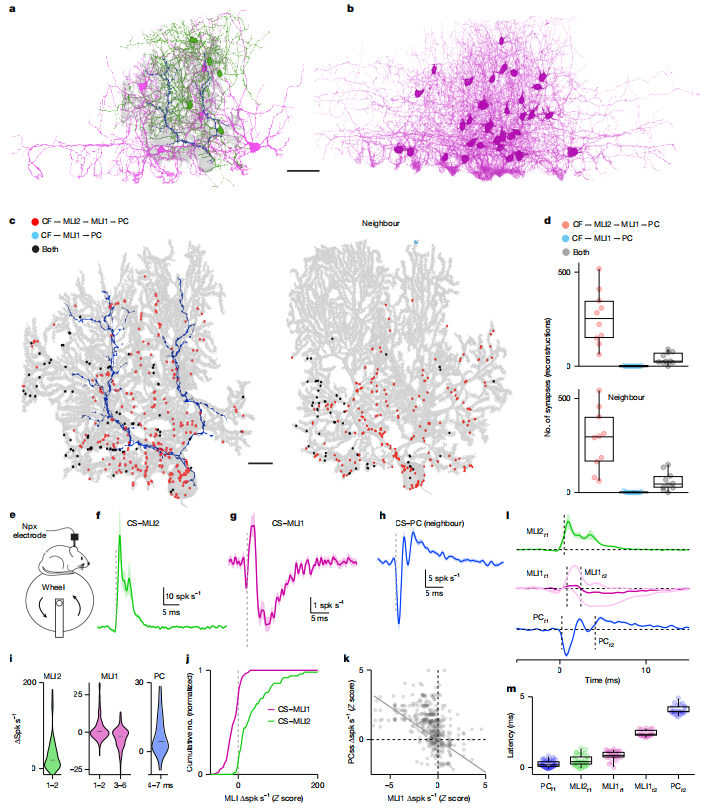
图3
4. 感觉输入下,MLI1的响应取决于CF与颗粒细胞的平衡
空气吹眼刺激同时激活:
· CF通路(去抑制)
· 苔藓纤维(MF)-颗粒细胞通路(直接兴奋MLI1)
当CF同步激活时,去抑制通路占优,MLI1被强烈抑制。
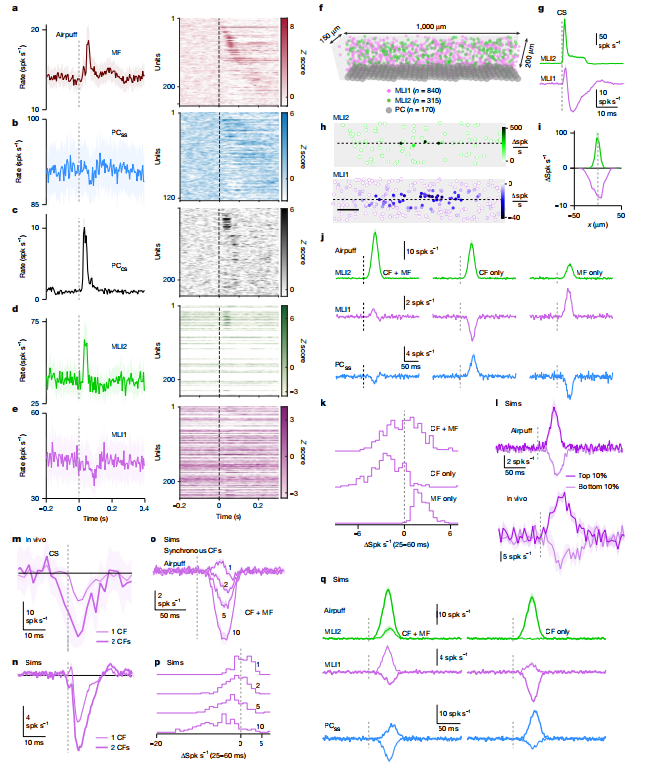
图4
5. CF同步性是增强PC钙信号的关键
双光子钙成像显示:
· 当邻近PC的CF同步激活时,PC树突钙信号显著增强;
· 这种增强效应在60 μm范围内最明显,且随同步CF数量增加而增强;
· 模拟实验证实,去除CF-MLI2连接后,去抑制效应消失,PC反而被抑制。
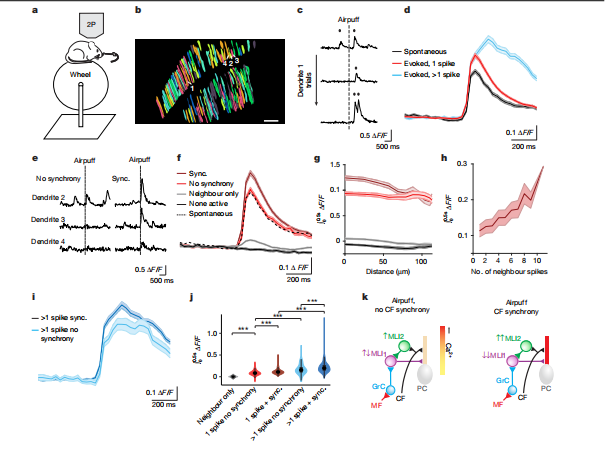
图5
研究意义与展望
理论意义:
· 为小脑“去抑制”环路提供了直接的解剖与功能证据;
· 解释了为何同步CF输入能高效诱导学习(如眨眼条件反射)。
未来方向:
· 特异性操控MLI2活性,验证其在学习中的必要性;
· 探索其他脑区是否存在类似“去抑制”微环路;
· 结合光遗传与钙成像,进一步解析CF同步性在行为学习中的动态作用。
总结:
攀缘纤维通过“谷氨酸溢出”优先激活去抑制性中间神经元MLI2,形成CF-MLI2-MLI1-PC通路,在同步激活时有效增强浦肯野细胞树突钙信号,为小脑运动学习提供关键机制支持。
邮箱:info@TianyaBio.com
电话:020-3107 8154
地址:广州市番禺区兴业大道东路1078号2-50
-

微信公众号
-

微信客服